








ThS. Nguyễn Tố Cát Triệu, Ths Nguyễn Thị Nhung
Trung tâm Nghiên cứu và Phát triển Cây hồ tiêu
1.Đặt vấn đề
Hồ tiêu là cây công nghiệp dài ngày, do đó yếu tố giống đóng vai trò rất quan trọng, quyết định đến hiệu quả sản xuất, kinh doanh trong suốt thời kỳ 20 – 25 năm. Một số giống phổ biến trong sản xuất hiện nay như: Vĩnh Linh, Lada, tiêu Sẻ, Ấn Độ, Lộc Ninh, Phú Quốc, tiêu Trâu… Trong đó, giống tiêu Vĩnh Linh là giống chủ lực, chiếm trên 90% diện tích. Trải qua một thời gian dài canh tác và nhân giống vô tính từ hom thân hoặc hom lươn, các giống tiêu có biểu hiện thoái hóa, suy giảm năng suất và khả năng chống chịu.
Để cải thiện đặc tính của các giống hồ tiêu nước ta thì phương pháp lai tạo giống là hết sức cần thiết. Bên cạnh đó, lai tạo giống còn tăng tính đa dang, phong phú về nguồn gen cây hồ tiêu. Tuy nhiên, để lai tạo giống thành công thì chúng ta cần phải hiểu rõ đặc điểm về hoa của cây hồ tiêu.
Bài viết này nhằm tổng hợp, phân tích các nghiên cứu trên thế giới về đặc điểm sinh lý hoa hồ tiêu, từ đó làm cơ sở khoa học cho việc thực hiện các nghiên cứu về lai hoa hồ tiêu ở Việt Nam.
2. Đặc điểm hình thái của hoa hồ tiêu
Cây hồ tiêu có tên khoa học là Piper nigrum L., thuộc họ Piperaceace, bộ Piperales, thuộc loại cây thân thảo.
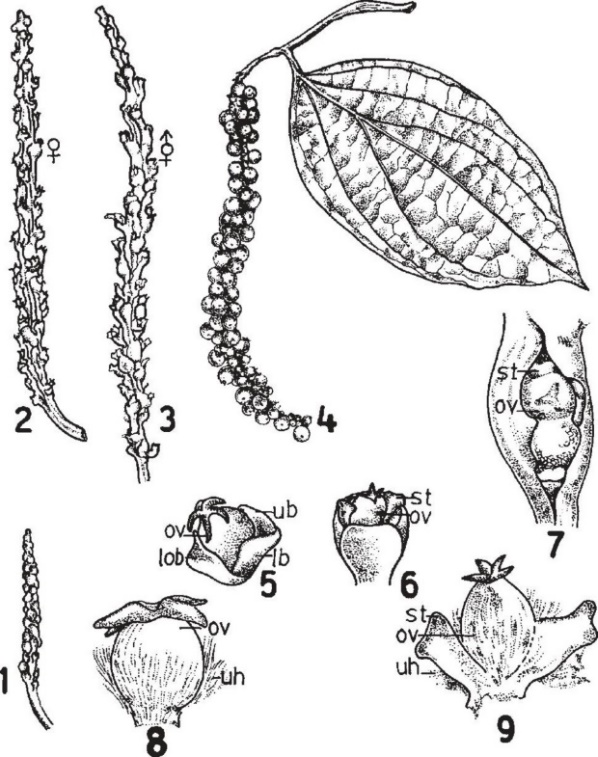
(Nguồn: Tucker, S.C. 1982)
Hình 1: Hình thái học của hoa tiêu
1.Gié tiêu non 2, 3.Gié tiêu trưởng thành 4.Gié quả 5. Hoa cái 6.Hoa lưỡng tính 7.Hai hoa nằm cạnh nhau và có lá bắc chung 8. Hoa cái 9.Hoa lưỡng tính. B: lá bắc, uh: lông tơ, ov: bầu nhụy, st: nhị, lob: lá bắc dưới cùng, lb: lá bắc dưới, ub: lá bắc trên
 (Nguồn: PRDC, 2017)
(Nguồn: PRDC, 2017)
Hình 2: Gié hoa tiêu chưa trưởng thành
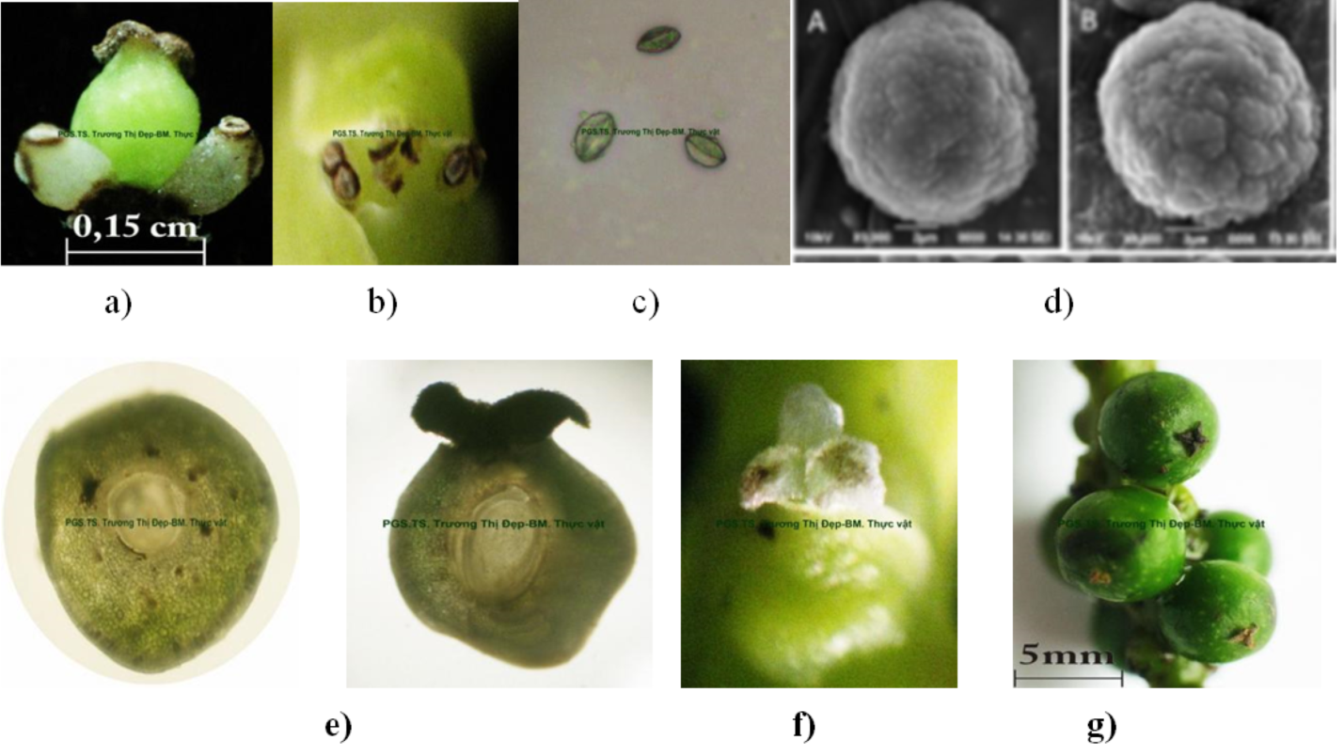
Hình 3: Cơ quan sinh sản hữu tính của cây hồ tiêu.
- – Hoa lưỡng tính
- – Nhị hoa
- – Bao phấn
- – Hạt phấn tiêu
- – Mặt cắt ngang và dọc của nhụy hoa ( noãn)
- – Đầu nhụy
- – Quả
Bộ nhị gồm 2 nhị nằm hai bên nhụy; chỉ nhị nạc dài 0,05 – 0,08 cm; mỗi nhị gồm 2 ô phấn, màu vàng nâu, dài 0,8 – 1 mm. Hạt phấn hình cầu, đường kính trung bình khoảng 8,84 – 11 µm (Rahiman, 1981).
Nhụy hoa tiêu gồm 3 – 4 lá noãn dính nhau tạo thành bầu trên 1 ô, đính đáy; bầu hình cầu màu xanh, đường kính 1 – 1,5 mm; vòi nhụy rất ngắn gần như không có; có 3 – 4 đầu nhụy dài 1 – 2 mm, màu trắng (Kanta, 1962; Tucker, 1982).
Bảng 1: Đặc tính của gié và quả một số giống
|
STT |
Giống |
Chiều dài gié (mm) |
Chiều dài cuống gié (mm) |
Hình dạng gié |
Dạng quả |
Kích thước quả |
|
1 |
Aimpiriyan |
115,6 |
12,2 |
Cong |
Tròn |
To |
|
2 |
Arakkulammunda |
114,0 |
13,1 |
Thẳng |
Tròn |
Trung bình |
|
3 |
Arimulahu |
80,4 |
13,2 |
Thẳng |
Tròn |
Nhỏ |
|
4 |
Balancotta |
127,4 |
17,2 |
Thẳng |
Tròn |
To |
|
5 |
Bilimalligesara |
105,6 |
8,6 |
Thẳng |
Tròn |
Trung bình |
|
6 |
Cheriyakaniakkadan |
105,4 |
9,2 |
Thẳng |
Trứng |
Nhỏ |
|
7 |
Cbeppuhulamundi |
120,3 |
19,0 |
Thẳng |
Tròn |
Trung bình |
|
8 |
Cholamundi |
111,3 |
6,4 |
Thẳng |
Tròn |
Nhỏ |
|
9 |
Jeerakmundi |
103,6 |
6,0 |
Thẳng |
Tròn |
Nhỏ |
|
10 |
Karimunda |
78,0 |
10,0 |
Thẳng |
Tròn |
Trung bình |
|
11 |
Kaniakkadan |
92,5 |
9,0 |
Thẳng |
Tròn |
Trung bình |
|
12 |
Kariuilanchy |
104,3 |
10,0 |
Thẳng |
Thuôn |
To |
|
13 |
Karimkotta |
156,3 |
21,0 |
Thẳng |
Tròn |
To |
|
14 |
Kalluually 1 |
69,2 |
12,0 |
Cong |
Tròn |
Nhỏ |
|
15 |
Kalluvally 2 |
124,9 |
10,6 |
Thẳng |
Tròn |
Trung bình |
|
16 |
Kallubalancotta |
136,4 |
10,5 |
Thẳng |
Tròn |
Trung bình |
|
17 |
Kottanadan |
106,9 |
11,1 |
Cong |
Tròn |
Trung bình |
|
18 |
Kucbing |
91,0 |
9,7 |
Cong |
Thuôn |
Trung bình |
|
19 |
Kuriyalmundi |
53,2 |
10,1 |
Cong |
Tròn |
Nhỏ |
|
20 |
Kuthiravally |
171,6 |
10,5 |
Thẳng |
Tròn |
Trung bình |
|
21 |
Kurimalai |
126,1 |
14,7 |
Thẳng |
Tròn |
Trung bình |
|
22 |
Malamundi |
96,7 |
7,9 |
Thẳng |
Tròn |
Trung bình |
|
23 |
Mundi |
86,3 |
9,1 |
Thẳng |
Tròn |
To |
|
24 |
Narayakkodi |
82,3 |
7,9 |
Cong |
Trứng |
Nhỏ |
|
25 |
Neelamundi |
97,0 |
7,4 |
Thẳng |
Tròn |
To |
|
26 |
Nedumchola |
51,7 |
9,6 |
Thẳng |
Trứng |
Nhỏ |
|
27 |
Neyyatinkaramundi |
71,0 |
7,0 |
Thẳng |
Tròn |
Nhỏ |
|
28 |
Ottaplackal 1 |
113,8 |
12,4 |
Thẳng |
Tròn |
Trung bình |
|
29 |
Panniyur 1 |
140,0 |
13,7 |
Thẳng |
Tròn |
To |
|
30 |
Perambramunda |
119,0 |
10,0 |
Thẳng |
Thuôn |
Trung bình |
|
31 |
Perumkodi |
117,6 |
13,1 |
Thẳng |
Tròn |
To |
|
32 |
Poonjaranmunda |
163,9 |
12,2 |
Thẳng |
Tròn |
To |
|
33 |
Sagar Local |
90,0 |
10,8 |
Thẳng |
Tròn |
To |
|
34 |
Theuanmundi |
96,5 |
8,7 |
Thẳng |
Thuôn |
Trung bình |
|
35 |
Tbommankodi |
127,6 |
15,5 |
Thẳng |
Tròn |
Trung bình |
|
36 |
Thulamundi |
95,0 |
10,6 |
Thẳng |
Tròn |
Trung bình |
|
37 |
Udakkere |
128,8 |
11,7 |
Thẳng |
Tròn |
To |
|
38 |
Uthirancotta |
105,6 |
12,6 |
Thẳng |
Tròn |
To |
|
39 |
Vadakkan |
117,3 |
14,6 |
Thẳng |
Tròn |
To |
|
40 |
Valiakanikkadan |
97,5 |
13,8 |
Thẳng |
Thuôn |
To |
3. Đặc điểm phân hóa và các giai đoạn phát triển của gié hoa hồ tiêu
Theo một số nghiên cứu ở Kerala – Ấn Độ, sự phân hóa của chồi xảy ra chủ yếu trong tháng 6 – 7, khi bắt đầu mùa mưa (K.A.U, 1984). Gồm 3 giai đoạn: (i) sự xuất hiện của mầm hoa hình nón bao quanh bởi lá bẹ (chưa khác biệt với mầm lá), (ii) sự kéo dài của chồi hoa (sự kéo dài của gié tiêu), (iii) Sự phân hóa của mô phân sinh bên (các mô nằm giữa tầng sinh bần và tầng sinh libe – gỗ). Sự phát triển của gié tiêu cũng có thể chia làm 5 giai đoạn: (i) hai mầm hình nón không khác biệt được bao bọc bởi lá bẹ, bắt đầu sự phân hóa của chồi, (ii) sau đó, một trong hai mầm nới rộng, kéo dài ra và một cấu trúc hình vòm xuất hiện ở đỉnh của cơ quan này, (iii) một cấu trúc dài giống như một cành tiêu xuất hiện, (iv) phân hóa các bộ phận của hoa, và (v) sự phân hóa được hoàn thành với sự xuất hiện của nhị và nhụy. Toàn bộ quá trình kéo dài khoảng 20 ngày.
4. Đặc điểm thụ phấn của hoa tiêu
– Điều kiện nở hoa: Vào thời kỳ ra hoa cây tiêu cần độ ẩm không khí lớn từ 70 – 90%, độ ẩm cao làm hạt phấn dễ dính vào nuốm nhụy và nuốm nhụy trương to kéo dài thời gian thụ phấn (Hoàng Thanh Tiệm và cs., 2007).
Hầu hết các giống tiêu, hoa sẽ nở theo hình thức lệch giao, nhụy sẽ nở trước nhị từ 1 – 14 ngày. Chỉ có một ít giống tiêu nhụy và nhị chín cùng lúc.
– Ở giai đoạn đầu của sự phát triển, gié và lá đối xứng được bao bọc trong lá bẹ, sau 3 – 4 tuần gié mới xuất hiện. Kế tiếp là sự xuất hiện của nhụy, thời gian xuất hiện khoảng từ 10 – 20 ngày phụ thuộc vào giống, một số giống nhụy xuất hiện sớm khoảng 5 ngày sau khi gié xuất hiện. Thời gian nhận phấn của nhụy từ 3 – 9 ngày, hoa nở trước có khoảng thời gian nhận phấn dài hơn hoa nở sau (K.A.U, 1978).
Ở điều kiện của vùng Sarawak – Malaysia, đối với giống P.Kuching, tính từ khi gié xuất hiện, nhụy xuất hiện sau 16 ngày, nhuỵ có khả năng tiếp nhận hạt phấn sau 22 ngày và bao phấn xuất hiện sau 27 ngày (Anon, 1979). Thành phần hoa trên gié khác nhau giữa các giống, có một số giống tất cả đều là hoa lưỡng tính như Karimunda, trong khi một số giống khác có cả hoa đơn tính như Panniyur 1 (97% hoa lưỡng tính, 3% hoa đơn tính) (IISR, 1997).
– Sự mở bao phấn: Quá trình này chịu ảnh hưởng bởi nhiệt độ và độ ẩm tương đối (Iljas 1960, Martin và Gregory, 1962). Ở Sarawak, Malaysia bao phấn hoa tiêu thường mở từ 12 giờ đến 14 giờ vào những ngày có độ ẩm tương đối 60% và nhiệt độ 32°C, kết hợp với ánh sáng mặt trời. Hầu hết phấn hoa phân tán tự do trên các nhụy lân cận và các phần khác của gié (De Waard và Zevan, 1969).
Nghiên cứu tại Sarawak, Malaysia cho thấy không có sự khác biệt có ý nghĩa về thời gian mở bao phấn ở các giống tiêu. Thông thường, sự phân tán hạt phấn trong điều kiện thực địa có thể thay đổi theo thời gian, phụ thuộc vào độ ẩm tương đối (RH) và nhiệt độ (Yates & Darrel, 1993). Ở giống Semongok Aman (SA), ở điều kiện đồng ruộng thời gian mở bao phấn trung bình từ 23 giờ – 24 giờ (Chen, C.S, 2018).
– Hạt phấn:
Số lượng hạt phấn: Các giống tiêu khác nhau có số lượng hạt phấn trong mỗi bao phấn khác nhau, ước tính mỗi gié có từ 100.000 – 300.000 hạt phấn (Martin và Gregory, 1962).
Tính hữu dục của hạt phấn: Kearns và Inouye (1993) cho rằng hạt phấn được thu ngay khi bao phấn mở là hữu dục nhất. Một nghiên cứu khác cho thấy rằng hạt phấn thu sau 20 giờ có tỷ lệ nảy mầm tương đối thấp và ống phấn ngắn. Phấn hoa thu được từ giai đoạn này có thể bất dục mặc dù đạt được tỷ lệ nảy mầm thích hợp và ống phấn dài. Nghiên cứu tính hữu dục của hạt phấn của 10 giống (Semongok Aman; Kuching; Semongok Emas; Semongok Perak; Semongok 1; Nyerigai; India; Lampung Daun Lebar; Sarikei; Yong Petai) trong 5 giai đoạn khi bao phấn nứt: khi bao phấn nứt, sau 5 giờ, 10 giờ, 15 giờ và 20 giờ cho thấy có sự khác biệt đáng kể về khả năng nảy mầm của hạt phấn, khả năng nảy mầm của hạt phấn cao nhất ở giai đoạn từ 0 – 15 giờ sau khi bao phấn nứt, sau 20 giờ khả năng nảy mầm của hạt phấn giảm đáng kể. Thời gian hữu dục của hạt phấn khác nhau giữa các giống (Chen, Y. S., Dayod, M. and Tawan, C. S., 2018).- Khả năng nhận phấn của đầu nhụy:
Nghiên cứu khả năng nhận phấn của đầu nhụy đối với 10 giống tiêu (Semongok Aman; Kuching; Semongok Emas; Semongok Perak; Semongok 1; Nyerigai; India; Lampung Daun Lebar; Sarikei; Yong Petai) trong 3 giai đoạn: bắt đầu xuất hiện đầu nhụy (ngày đầu tiên); giai đoạn đầu nhụy mở ra (đầu nhụy xuất hiện ngày 2 – 3) và đầu nhụy mở hoàn toàn (đầu nhụy xuất hiện ngày thứ 4 – 6) cho thấy các giống có cùng xu hướng tiếp nhận. Đầu nhụy ở giai đoạn 1 có mức độ tiếp nhận thấp hơn đáng kể so với giai đoạn 2, 3. Purseglove (1968) cho rằng đầu nhụy có khả năng tiếp nhận tốt nhất sau 3 – 5 ngày xuất hiện và có khả năng nhận phấn lên tới 10 ngày. Tuy nhiên, nghiên cứu của Chen (2011) lại cho thấy không có sự khác biệt đáng kể về khả năng tiếp nhận của đầu nhụy ở cả 3 giai đoạn.
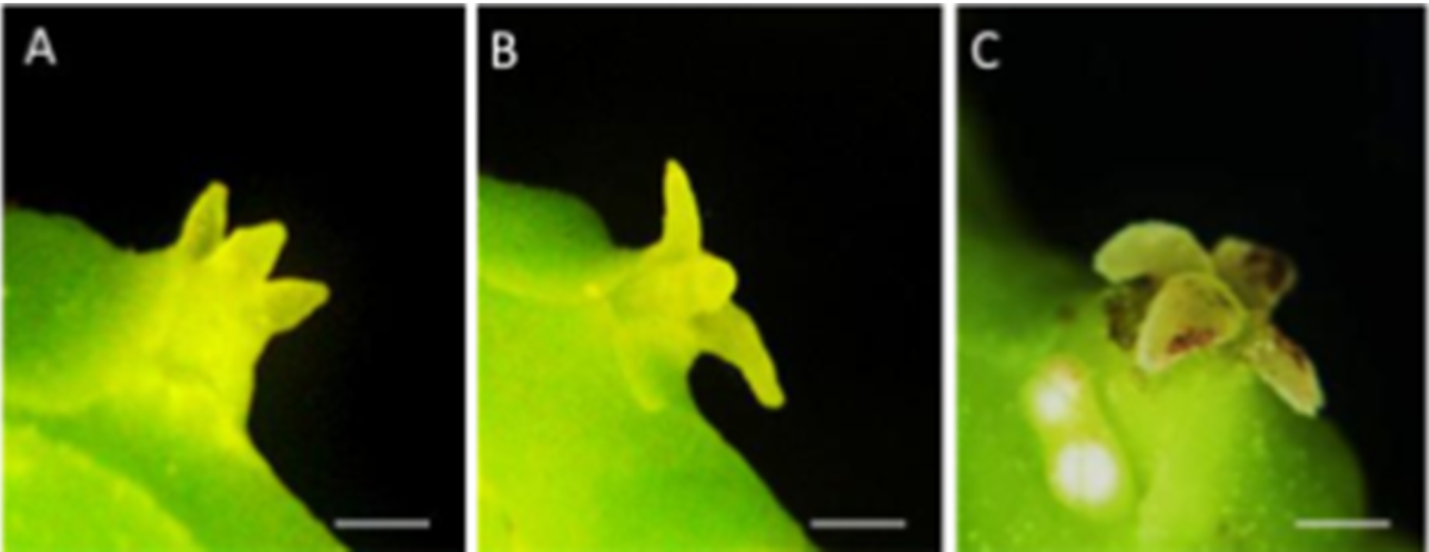
Nguồn: Helen, J. Y., & Lauren, G. (2002)
Hình 4: A:Giai đoạn 1, sự xuất hiện đầu tiên của đầu nhụy. B: Giai đoạn 2, đầu nhụy mở ra. C: Giai đoạn 3, đầu nhụy mở hoàn toàn.
Helen và Lauren (2002) cho rằng thời gian xuất hiện của đầu nhụy không ảnh hương tới khả năng tiếp nhận hạt phấn. Đầu nhụy có khả năng tiếp nhận ở bất kỳ giai đoạn nào trước khi nhận phấn. Sau khi đầu nhụy nhận phấn, các tế bào đầu nhụy khô héo, đồng thời các hạt phấn hydrat hóa và nảy mầm (Wetzstein & Sparks, 1989). Nếu không được thụ phấn, đầu nhụy có thể vẫn tiếp tục tồn tại trong một thời gian lâu hơn (Wetzstein & Sparks, 1989).
– Phương thức thụ phấn: Cây hồ tiêu là cây tự thụ phấn. Phấn của hoa này sẽ được thụ cho hoa khác trên cùng 1 cây (Geitonogamy). Hoa tiêu có đặc điểm cấu trúc để thích nghi với phương thức thụ phấn này như: gié đơn độc, hoa sắp xếp trình tự theo kiểu xoắn ốc, hiện tượng lệch giao… Iljas (1960) phát hiện ra rằng các gié được bọc trong túi polyethylene vẫn cho bộ quả tốt không phụ thuộc vào côn trùng hay nước mưa.
Hoa thụ phấn nhờ vào ẩm độ không khí. Độ ẩm không khí cao thuận lợi cho sự phân tán hạt phấn ra khỏi bao phấn và giúp vòi nhụy giữ sự cương được lâu để dễ dàng tiếp nhận hạt phấn (Phan Hữu Trinh et al., 1988; Phan Quốc Sủng, 2000). Hạt phấn mang nhiều chất dính, trong trường hợp mưa nhẹ sẽ làm vỡ những hạt chất dính của hạt phấn và làm cho hạt phấn bám vào chất nhầy của núm nhụy do đó đã gia tăng hiệu quả của sự phân bố hạt phấn. Martin và Gregory (1962) dựa trên nghiên cứu của họ ở Puerto Rico đã chỉ ra rằng 32 – 64% phấn hoa trên gié có thể được phân tán trong không khí trong vòng 24 giờ sau khi bung phấn.
5. Kết luận
Như vậy, sự lệch giao chính là đặc điểm khác biệt nhất của các giống tiêu, trên cùng một hoa lưỡng tính nhụy xuất hiện trước, sau đó nhụy mở ra và nhận phấn. Nhị của chính hoa đó sẽ chín sau đó, bao phấn mở ra và thực hiện quá trình thụ phấn, thụ tinh. Căn cứ vào đặc điểm này có thể thực hiện việc thụ phấn nhân tạo giữa các giống tiêu, tuy nhiên phải thực hiện kỹ thuật khử đực bằng cách loại bỏ bao phấn.
Tài liệu tham khảo
Tiếng Việt
- Hoàng Thanh Tiệm, Lê Ngọc Báu, Tôn Nữ Tuấn Nam, Nguyễn Hùng, Nguyễn Thị Thoa (2007), Kỹ thuật trồng, chăm sóc và chế biến hồ tiêu, Nhà xuất bản Nông Nghiệp Hà Nội.
- Nguyễn Mạnh Chinh, Nguyễn Đăng Nghĩa (2007), ”Trồng – chăm sóc & phòng trừ sâu bệnh hồ tiêu”, Bác sĩ cây trồng quyển 22, nhà xuất bản nông nghiệp thành phố Hồ Chí Minh.
- Phan Hữu Trinh và ctv (1988). Kỹ Thuật Trồng Tiêu. Nhà Xuất Bản Nông Nghiệp.
4. Phan Quốc Sủng (2000). Tìm Hiểu về Kỹ Thuật Trồng và Chăm Sóc Cây Hồ Tiêu. Nhà Xuất Bản Nông Nghiệp
Tiếng Anh
- Chen, Y. S. (2011). A study on interspecific hybridization between Piper nigrum and Piper colubrinum. (Unpublished Master thesis). Universiti Malaysia Sarawak, Kota Samarahan, Sarawak, Malaysia.
- Chen, Y. S., Dayod, M. and Tawan, C. S. (2018). Anther Dehiscence, Pollen Viability and Stigma Receptivity Study on Cultivars of Black Pepper (Piper nigrum). Tropical Agricultural Science, 41 (2), 801 – 814.
- Chen, Y. S., Tawan. C. S., Paulus, A. D., Ng, S. C., & Sim, S. L. (2010). Developing method of interspecific hybridization between Piper nigrum and Piper colubrinum. In D. Kanakaruju, K. Meekiong, I. B. Ipor, C. K. Lim, P. T. Lim, H. A. Roslan (Eds.), Proceedings of the 3rd Regional Conference on Natural Resources in the Tropics (pp. 293-300). Kota Samarahan, Sarawak: Universiti Malaysia Sarawak.
- Helen, J. Y., & Lauren, G. (2002). Reproductive Biology: The effects of stigma age on receptivity in Silene alba (Caryophyllaceae). American Journal of Botany, 89(8), 1237-1241.
- IISR (1997) Ann. Rep. 1996-1997, Indian Institute of Spices Research, Calicut.
- A.U. (1984) Research Report (1982–83). Kerala Agri. Uni., Vellanikkara, Thrissur, pp. 182– 185.
- Kearns, C. A., & Inouye, D. W. (1993). Techniques for pollination biologists. American Journal of Botany, 94, 1183-1192.
- Martin, F.W. and Gregory, L.E. (1962) Mode of pollination and factors affecting fruit set of Piper nigrum L. in Puerto Rico. Crop Sci., 5; 295–299.
- Tucker, S.C. (1982). Inflorescence and flower development in the Piperaceae. II. Inflorescence development of Piper. Amer J. Bot., 69, 1389–1401.
- Wetzstein, H. Y., & Sparks, D. (1989). Stigma-Pollen interactions in pecan. Journal America Society Horticulture Science, 114, 355-359.
- Yates, I. E., & Darrel, S. (1993). Environmental regulation of anther dehiscence and pollen germination in Pecan. Journal of America Society of Horticulture Science, 118(6), 699-706.